


Estudios de investigación anteriores sugieren que el L-aspartato, al igual que el L-glutamato, desencadena actividad excitatoria en las neuronas. El L-aspartato funciona con el L-glutamato en las vesículas sinápticas de las sinapsis excitatorias asimétricas. Pero, la concentración total de estos en el cerebro humano (0.96-1.62 μmol/gramo de peso húmedo), sus concentraciones extracelulares en la corteza medida por microdiálisis (1.62 μM para L-aspartato y 9.06 μM para L-glutamato) y su suministro según la inmunohistoquímica sugieren que el L-aspartato es significativamente menos abundante que el L-glutamato. Además, el L-aspartato es un potente agonista de los receptores NMDA pero no de otros iGluR con una CE50 solo ocho veces mayor que la del L-glutamato. Los EAAT, que desempeñan un papel fundamental en la captación de todo el L-glutamato liberado por las vesículas en el sistema nervioso central (SNC), también requieren la utilización de L-aspartato. El L-aspartato es quizás tan menos esencial como el L-glutamato relacionado con la actividad excitadora total asociada con los iGluR. Junto con su papel como neurotransmisor, como se mencionó anteriormente, el L-aspartato también es necesario como sustrato para la aspartato aminotransferasa, que se convierte en 2-oxoglutarato y L-glutamato para transportarse a las vesículas corticales de las neuronas glutamatérgicas que, en consecuencia, también pueden aumentar indirectamente la liberación de L-glutamato. UN
Otras moléculas en la señalización de glutamato
Una característica que distingue a los receptores NMDA de diferentes iGluR es que la activación de los receptores NMDA necesita la conexión de un coagonista a la región de unión a glicina del receptor. Por ejemplo, en la retina y en la médula espinal, el origen de la glicina puede desbordarse de las sinapsis inhibitorias glicinérgicas. Pero, en diferentes regiones del cerebro con aumento de la expresión del receptor NMDA, como la formación del hipocampo, faltan reacciones asociadas con los receptores de glicina sensibles a la estricnina, al menos en las neuronas adultas, lo que demuestra la ausencia de neurotransmisiones inhibitorias glicinérgicas. Sin embargo, la glicina se encuentra en el líquido extracelular del hipocampo en cantidades iniciales de aproximadamente 1.5 μM, que es similar a la saturación de la región de unión de glicina del receptor NMDA, aunque estos pueden estar regulados hacia arriba y hacia abajo. El origen de la glicina extracelular en el hipocampo pueden ser las neuronas que liberan glicina a través del transportador de aminoácidos alanina-serina-cisteína 1 (asc-1). Pero también se ha demostrado la liberación de glicina por los astrocitos que es estimulada por la despolarización y el kainato. Se requieren más estudios de investigación para finalmente mostrar estas medidas de resultado. UN
Incluso en estudios de investigación anteriores del receptor NMDA y su coactivación por la glicina, se reveló que los D-aminoácidos, particularmente la D-serina, son casi tan poderosos como la glicina. Solo varios años después, se hizo evidente que la D-serina se encuentra en cerebros humanos y de ratas en aproximadamente un tercio de su concentración de L-serina, que tiene una concentración absoluta de más de 0.2 μmol/g de tejido cerebral. Utilizando un antisuero para D-serina, los estudios de investigación demostraron que la D-serina del cerebro solo se encuentra en los astrocitos y su suministro se ajusta a la expresión de los receptores NMDA. Además, los mismos investigadores demostraron que los astrocitos cultivados liberan D-serina cuando se exponen a L-glutamato o kainato. La abundancia de D-serina la encuentra la enzima degradante D-aminoácido oxidasa (DAO) que revela una mayor expresión en el cerebro posterior donde se reducen los niveles de D-serina, así como la enzima sintética serina racemasa que crea D-serina a partir de L- serina La D-serina parece almacenarse en vesículas citoplasmáticas en los astrocitos y puede liberarse por exocitosis. La potenciación a largo plazo depende de la liberación de D-serina de los astrocitos en cortes de hipocampo, lo que sugiere que este aminoácido definitivamente juega un papel fundamental en la neurotransmisión glutamatérgica a través de los receptores NMDA. Además, en cortes de hipocampo, los estudios encontraron, utilizando enzimas degradantes de D-serina y glicina, que la D-serina funciona como un cotransmisor para los receptores NMDA sinápticos en las neuronas CA1 y que la glicina funciona como el co-agonista endógeno para los receptores NMDA extrasinápticos. Los receptores NMDA sinápticos de las neuronas de la circunvolución dentada utilizan glicina en lugar de D-serina como coagonista. UN
En conjunto, las medidas de resultados de varios niveles muestran que el L-aspartato no funciona simplemente como un agonista de los receptores NMDA, sino que también la glicina y la D-serina desempeñan un papel fundamental en la neurotransmisión glutamatérgica en el cerebro humano. Pero también se ha demostrado que otras moléculas son moduladores relevantes de la neurotransmisión glutamatérgica. UN
Glutamato Activado por Otras Moléculas
L-homocisteato (L-HCA) tiene similitudes estructurales con L-glutamato. El aminoácido no proteico es un producto de oxidación de la homocisteína que se biosintetiza a partir de la metionina en la eliminación de su propio grupo metilo terminal y también es un intermediario de la ruta de transulfuración por la cual la metionina se puede convertir en cisteína a través de la cistationina. Los primeros estudios de investigación demostraron que este aminoácido puede causar la entrada de calcio en las neuronas cultivadas de forma tan segura y eficaz como el L-glutamato. Además, L-HCA reveló una mayor afinidad por los receptores NMDA en comparación con otros iGluR en ensayos de unión asociados con su capacidad para causar excitotoxicidad inhibible por antagonistas del receptor NMDA y afluencia de sodio. Además, L-HCA puede desencadenar mGluR5 tan eficientemente como L-glutamato. L-HCA se encuentra en el cerebro, sin embargo, se demostró que las concentraciones son aproximadamente 500 veces menores que las de L-glutamato e incluso 100 veces menores en comparación con las de L-aspartato en diferentes regiones del cerebro de rata. Durante la estimulación inducida por potasio, la descarga de L-HCA se activa a partir de preparaciones de cortes de cerebro, como se demostró con L-aspartato y L-glutamato, aunque la liberación absoluta de HCA es aproximadamente 50 veces menor. Sorprendentemente, HCA es un inhibidor competitivo muy eficaz de la absorción de cistina y L-glutamato a través del sistema antiportador de cistina/glutamato xâˆ'c, la actividad que regula y gestiona las concentraciones de L-glutamato extrasináptico extracelular en el cerebro. Por lo tanto, el impacto de L-HCA en la activación de NMDA y otros receptores de L-glutamato también puede depender del desencadenante de L-glutamato inducido por L-HCA a través del sistema xâˆ'c. L-HCA puede desempeñar un papel importante en la estimulación general de los receptores de L-glutamato. Sin embargo, esto puede cambiar enormemente bajo ciertas condiciones, por ejemplo, en pacientes con terapia de dosis altas de metotrexato, un medicamento contra el cáncer que, al restringir la dihidrofolato reductasa, limita el reciclaje de metionina a partir de homocisteína catalizada por tetrahidrofolato. Aquí, se demostraron concentraciones de L-HCA de más de 100 μM en el líquido cefalorraquídeo, mientras que L-HCA fue indetectable en los sujetos de control. Aún se requieren más estudios de investigación para determinar estas medidas de resultado. UN
Otras pequeñas moléculas endógenas que se cree que afectan la señalización del L-glutamato incluyen varios intermediarios del metabolismo del triptófano, como se muestra en la Figura 2. A través de la actividad de la indolamina 2,3-dioxigenasa (IDO) o triptófano 2,3-dioxigenasa (TDO), el triptófano se convierte en N-formil-L-quinurenina, que luego se convierte en quinurenina (KYN) por la formamidasa. Tres vías, dos de las cuales se conectan en un paso posterior, dan como resultado un mayor metabolismo. Primero, a través de la actividad de la quinurenina aminotransferasa (KAT), KYN se convierte en ácido quinurénico (KYNA). KYN también se puede convertir en 3-hidroxiquinurenina (3HK) mediante la quinurenina monooxigenasa (KMO), que posteriormente puede ser utilizada como sustrato por la quinureninasa para la síntesis del ácido 3-hidroxiantranílico (3HANA). Además, al utilizar KYN como sustrato, la quinureninasa desarrolla ácido antranílico (ANA), que por hidroxilación no específica también puede convertirse en 3HANA. Según estudios de investigación, 3HANA finalmente funciona como sustrato para la generación de ácido quinolínico (QUIN). UN
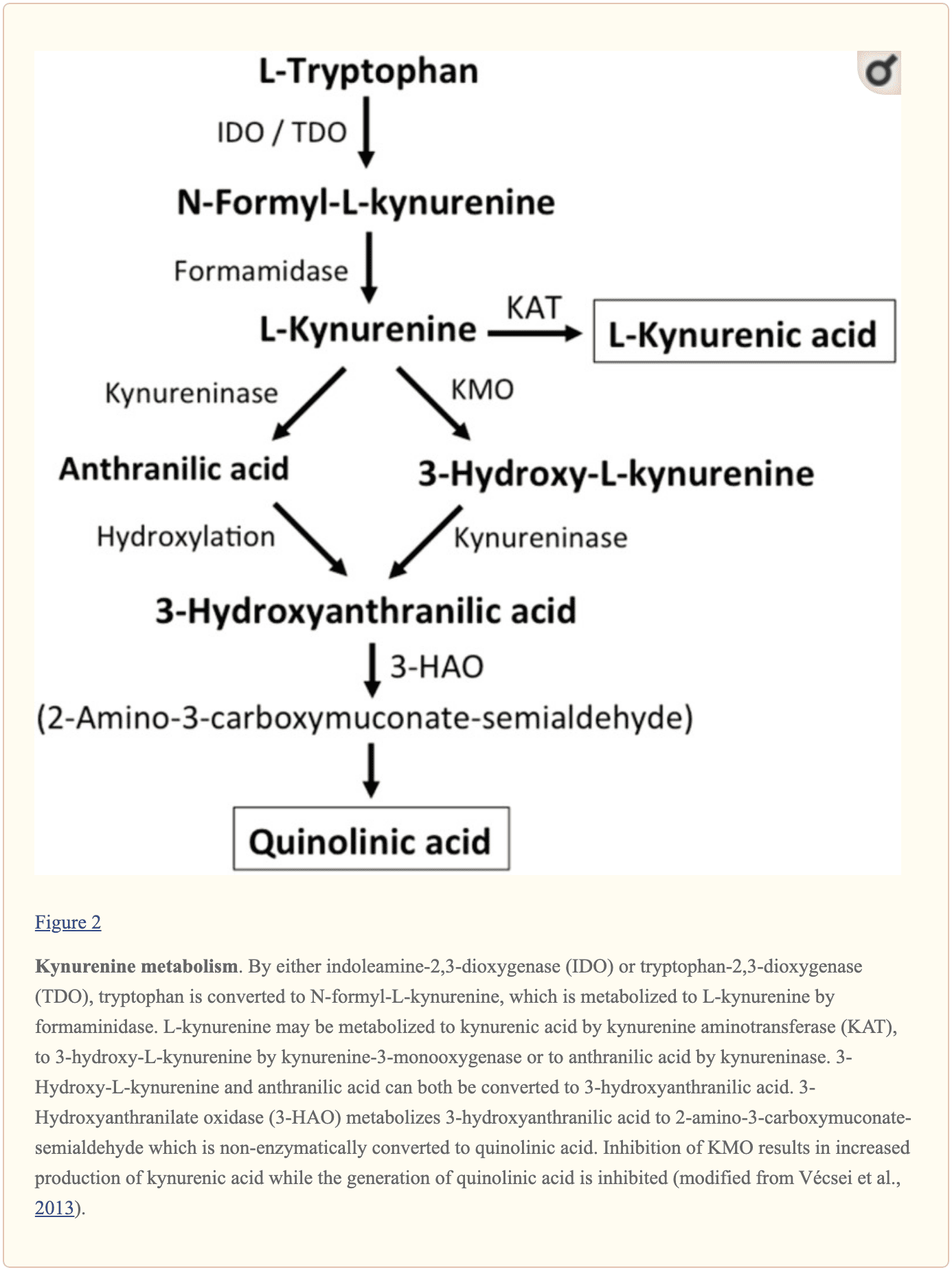
La concentración de triptófano en el cerebro de la rata es de aproximadamente 25 nmol/g de peso húmedo y aproximadamente 400 veces menor que la del L-glutamato y 100 veces menor que la del L-aspartato. Los niveles cerebrales demostrados de quinureninas son aún más bajos con 0.4-1.6 nmol/g para QUIN, 0.01-0.07 nmol/ml para KYNA y 0.016 nmol/g para 3HANA. Aproximadamente el 40 por ciento del cerebro KYN se sintetiza localmente. Los metabolitos del triptófano muestran una unión diferencial a las proteínas plasmáticas y su transporte a través de la barrera, que es bastante diferente. KYN y 3HK se transportan a través del gran sistema de transporte de aminoácidos neutros L. Las quinureninas parecen penetrar en el cerebro humano por difusión pasiva. Además, KYNA, 3HANA y, especialmente, ANA se unen a proteínas séricas que finalmente restringen y limitan su difusión a través de la barrera hematoencefálica. UN
Los estudios de investigación demostraron que QUIN, cuando se utilizaba ionoforéticamente en células de rata, provocaba disparos neuronales que habían sido prevenidos por un antagonista del receptor NMDA, lo que sugiere que QUIN podría funcionar como un agonista del receptor NMDA. Sin embargo, se ha demostrado que la EC50 de QUIN para activar las corrientes del receptor NMDA es aproximadamente 1000 veces mayor que la EC50 de L-glutamato. Se demostró que la inyección intracerebral de QUIN causa cambios ultraestructurales, neuroquímicos y de comportamiento similares a los causados por los agonistas del receptor NMDA. El hecho de que las concentraciones de QUIN sean entre 5000 y 15,000 61 veces más bajas que las concentraciones cerebrales de L-glutamato hace que sea poco probable que la modulación de la señalización del receptor NMDA por parte de QUIN desempeñe un papel esencial. Se demostró que KYNA funciona como un antagonista del receptor NMDA. Pero, aunque la infusión con el inhibidor de KMO Ro 8048-10 mejoró 6 veces las concentraciones de KYNA extracelular cerebral, esto no resultó en una inhibición de la despolarización neuronal mediada por NMDA, un hallazgo que desafía la creencia de que KYNA en cantidades casi fisiológicas directamente modula los receptores NMDA. En comparación, el aumento de KYNA en el cerebro inducido por el inhibidor de KMO JM35 disminuyó la concentración de L-glutamato cerebral extracelular. Además, los niveles de KYNA del líquido cerebral extracelular se han asociado con los niveles de L-glutamato, lo que sugiere que incluso a niveles fisiológicos o casi fisiológicos, KYNA modula el metabolismo del L-glutamato. Tanto la activación del receptor acoplado a proteína G GPR7 como la inhibición de los receptores nicotínicos de acetilcolina αXNUMX presinápticos se sugieren en la reducción inducida por KYNA en la liberación de L-glutamato. En resumen, aunque QUIN y L-HCA están presentes en el cerebro humano, sus concentraciones discuten en su contra con funciones en la regulación y mantenimiento de la neurotransmisión. Por el contrario, aunque las vías deben definirse con mayor detalle, la evidencia respalda los niveles y la opinión de que la descarga puede ser modulada por KYNA y la neurotransmisión. UN

El glutamato, junto con el aspartato y otras moléculas, son varios de los principales neurotransmisores excitadores del cerebro humano. Si bien estos desempeñan un papel fundamental en la estructura y función general del sistema nervioso central, incluido el cerebro y la médula espinal, las cantidades excesivas de otras moléculas pueden desencadenar receptores de glutamato. El exceso de glutamato puede causar excitotoxicidad que puede conducir a una variedad de problemas de salud, como la enfermedad de Alzheimer y otros tipos de enfermedades neurológicas. El siguiente artículo describe cómo otras moléculas pueden activar los receptores de glutamato. - Dr. Alex Jiménez DC, CCST Insight - Dr. Alex Jimenez DC, CCST Insight
Tratamiento de neuropatía con LLLT
Los estudios de investigación sugieren que el L-aspartato, como el L-glutamato, desencadena la actividad excitadora. L-aspartato funciona con L-glutamato en las vesículas sinápticas de sinapsis excitadoras asimétricas. Pero, la concentración total de estos en el cerebro humano sugiere que el L-aspartato es significativamente menos abundante que el L-glutamato. Además, el L-aspartato es un poderoso agonista para los receptores NMDA pero no para otros iGluR con un EC50 solo ocho veces mayor que el del L-glutamato. El alcance de nuestra información se limita a cuestiones de salud quiropráctica, musculoesquelética y nerviosa, así como a artículos, temas y debates sobre medicina funcional. Utilizamos protocolos funcionales de salud para tratar lesiones o trastornos crónicos del sistema musculoesquelético. Para seguir discutiendo el tema anterior, no dude en preguntarle al Dr. Alex Jiménez o contáctenos en 915-850-0900 . UNA
Curada por el Dr. Alex Jimenez Â
Referencias Â
- Lewerenz, Jan y Pamela Maher. "Toxicidad crónica del glutamato en enfermedades neurodegenerativas: ¿cuál es la evidencia?" Fronteras en Neurociencias, Frontiers Media SA, 16 Dec. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4679930/.
Discusión de tema adicional: dolor crónico
El dolor repentino es una respuesta natural del sistema nervioso que ayuda a demostrar posibles lesiones. A modo de ejemplo, las señales de dolor viajan desde una región lesionada a través de los nervios y la médula espinal hasta el cerebro. El dolor generalmente es menos intenso a medida que la lesión se cura, sin embargo, el dolor crónico es diferente al tipo promedio de dolor. Con dolor crónico, el cuerpo humano continuará enviando señales de dolor al cerebro, independientemente de si la lesión se ha curado. El dolor crónico puede durar varias semanas o incluso varios años. El dolor crónico puede afectar enormemente la movilidad de un paciente y puede reducir la flexibilidad, la fuerza y la resistencia.
Neural Zoomer Plus para Enfermedades Neurológicas
 Â
Â
El Dr. Alex Jiménez utiliza una serie de pruebas para ayudar a evaluar enfermedades neurológicas. El zoomero neuralTM Plus es un conjunto de autoanticuerpos neurológicos que ofrece reconocimiento específico de anticuerpo a antígeno. El zoomero neuronal vibranteTM Plus está diseñado para evaluar la reactividad de un individuo a 48 antígenos neurológicos con conexiones a una variedad de enfermedades relacionadas neurológicamente. El Zoomer Neural VibranteTM Plus tiene como objetivo reducir las condiciones neurológicas al brindar a los pacientes y médicos un recurso vital para la detección temprana de riesgos y un enfoque mejorado en la prevención primaria personalizada. UN
Fórmulas para soporte de metilación

de XYMOGEN Las fórmulas profesionales exclusivas están disponibles a través de profesionales de atención médica con licencia seleccionados. La venta por internet y el descuento de fórmulas XYMOGEN están estrictamente prohibidos.
Orgullosamente, El Dr. Alexander Jimenez hace que las fórmulas de XYMOGEN estén disponibles solo para los pacientes bajo nuestro cuidado.
Llame a nuestro consultorio para que podamos asignar una consulta médica para acceso inmediato.
Si eres paciente de Clínica médica de lesiones y quiropráctica, puedes preguntar por XYMOGEN llamando 915-850-0900.
![]()
Para su conveniencia y revisión de la XYMOGEN productos por favor revise el siguiente enlace. *XYMOGEN-Catalog-Descargar Â
* Todas las políticas anteriores de XYMOGEN permanecen estrictamente en vigor.
Â
Publicar descargos de responsabilidad
Alcance de la práctica profesional *
La información aquí contenida en "Neurología funcional: otras moléculas en glutamato" no pretende reemplazar una relación personal con un profesional de la salud calificado o un médico con licencia y no es un consejo médico. Lo alentamos a que tome decisiones de atención médica basadas en su investigación y asociación con un profesional de la salud calificado.
Información del blog y debates sobre el alcance
Nuestro alcance informativo se limita a la quiropráctica, musculoesquelética, medicina física, bienestar, contribuyendo etiológico alteraciones viscerosomáticas dentro de las presentaciones clínicas, la dinámica clínica del reflejo somatovisceral asociado, los complejos de subluxación, los problemas de salud delicados y/o los artículos, temas y debates de medicina funcional.
Brindamos y presentamos colaboración clínica con especialistas de diversas disciplinas. Cada especialista se rige por su ámbito de práctica profesional y su jurisdicción de licencia. Utilizamos protocolos funcionales de salud y bienestar para tratar y apoyar la atención de lesiones o trastornos del sistema musculoesquelético.
Nuestros videos, publicaciones, temas, asuntos e ideas cubren cuestiones clínicas, problemas y temas que se relacionan y respaldan directa o indirectamente nuestro ámbito de práctica clínica.*
Nuestra oficina ha intentado razonablemente proporcionar citas de apoyo y ha identificado el estudio o los estudios de investigación relevantes que respaldan nuestras publicaciones. Proporcionamos copias de los estudios de investigación de respaldo disponibles para las juntas reguladoras y el público a pedido.
Entendemos que cubrimos asuntos que requieren una explicación adicional de cómo puede ayudar en un plan de atención o protocolo de tratamiento en particular; por lo tanto, para discutir más a fondo el tema anterior, no dude en preguntar Dr. Alex Jiménez, DC, o póngase en contacto con nosotros en 915-850-0900.
Estamos aquí para ayudarlo a usted y a su familia.
Bendiciones
El Dr. Alex Jimenez corriente continua MSACP, enfermero*, CCCT, IFMCP*, CIFM*, ATN*
email: coach@elpasomedicinafuncional.com
Licenciado como Doctor en Quiropráctica (DC) en Texas & New Mexico*
Número de licencia de Texas DC TX5807, Nuevo México DC Número de licencia NM-DC2182
Licenciada como Enfermera Registrada (RN*) en Florida
Licencia de Florida N.° de licencia de RN RN9617241 (Control No. 3558029)
Estado compacto: Licencia multiestatal: Autorizado para ejercer en 40 Estados*
Matriculado actualmente: ICHS: MSN* FNP (Programa de enfermera practicante familiar)
Dr. Alex Jiménez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Mi tarjeta de presentación digital




 De nuevo te doy la bienvenida.
De nuevo te doy la bienvenida.
Los comentarios están cerrados.